Número 011 2002 Revista Internacional de Psicoanálisis Aperturas
Neurobiología de las interacciones estrés-memoria
Autor: de Iceta Ibáñez de Gauna, MarianoPalabras clave
Diamond, Estres, Kim, Hipocampo, Hormonas, Memoria, Neurobiologia.
- Artículo: Kim, J.J.; Diamond, D.M. The stressed hippocampus, synaptic plasticity and lost memories. Nature Reviews: Neuroscience, 2002, vol. 3: 453-62.
- Autor de la reseña: Mariano de Iceta
Nota introductoria: Es conocido a través de múltiples trabajos como el estrés afecta anatómicamente al hipocampo (un área cerebral de particular importancia para la memoria y un enclave central de la regulación neuroendocrina del organismo, y fisiológicamente a la memoria) (Kim & Yoon, 1998; McEwen, 2000). Asimismo, existen numerosas evidencias que dicha alteración está mediada en buena medida por los corticoides (Newcomer et al., 1994).
El trabajo que se reseña aborda en profundidad las relaciones entre el estrés y la memoria y, más allá de su densidad en cuanto a detalles técnicos, aporta, desde mi punto de vista, dos aspectos de particular interés: Primero, una definición del estrés que tiene en cuenta la controlabilidad del estímulo. Y segundo, plantea un modelo integrado que abarca la interacción entre las vías neurales y las hormonales (corticoides y otros neuromoduladores) en la génesis de las alteraciones en la memoria debidas al estrés.
El estrés, como factor ambiental que contribuye al deterioro de la memoria, constituye un problema significativo en la sociedad actual, con un envejecimiento e incremento en la población crecientes. El término estrés1 proviene de la ingeniería y, en su aplicación a lo biológico, se define como cualquier condición que perturba seriamente la homeostasis fisiológica/psicológica de un organismo. En las décadas recientes, numerosos estudios en neurociencia han mostrado que las experiencias estresantes pueden tener un impacto negativo en ciertas funciones cerebrales (deterioro en las capacidades de aprendizaje y de memoria, incremento del detrioro cognitivo relacionado con la edad, aumento de la susceptibilidad de las neuronas del hipocampo a sufrir atrofia o necrosis en respuesta a demandas metabólicas). No obstante, conviene recordar que la respuesta aguda al estrés (por ejemplo, incremento de la atención) se considera un mecanismo adaptativo que capacita al organismo para responder de manera efectiva a una amenaza, potencial o real, para su supervivencia.
La relación entre el estrés y la memoria es compleja. Si bien las experiencias estresantes, sobre todo las traumáticas, producen intensos y vívidos recuerdos que perduran en el tiempo de la situación traumática; no es menos cierto que el estrés deteriora la atención posterior y la memoria, llegando incluso a inducir una amnesia profunda.
Estrés y memoria hipocampal
El hipocampo, situado en la parte medial del lóbulo temporal, constituye una estructura cerebral que juega un papel crucial tanto en la memoria como en la regulación neuroendocrina de las hormonas del estrés. Es necesario para la formación de memoria declarativa (explícita) estable en humanos, y el equivalente en roedores (memoria espacial, o relacional/contextual). Es, asímismo, una de las dianas de las hormonas del estrés, con una de las mayores concentraciones de receptores para corticosteroides del cerebro de los mamíferos. Una función neuroendocrina del hipocampo es participar en la terminación de la respuesta de estrés por medio de una retroalimentación negativa que inhibe el eje hipotálamo-hipofiso-adrenal (Eje HHA).
En el hipocampo existen dos tipos de receptores para corticosteroides, los mineralocorticoides (MRs) y los glucocorticoides (GRs). Estos últimos, de menor afinidad y ocupados de manera masiva por corticosteroides en la respuesta al estrés, son los principales mediadores de los efectos adversos del estrés en el hipocampo (afectan negativamente al metabolismo neuronal, la supervivencia celular, las funciones fisiológicas y la morfología neuronal del hipocampo en las ratas). Numerosos estudios en las dos últimas décadas apoyan la idea de que el estrés y las hormonas del estrés deterioran las formas de memoria dependientes del hipocampo, tanto en humanos como en animales. Así por ejemplo, los pacientes con trastorno por estrés postraumático (TEP) presentan atrofia del hipocampo y déficits marcados en las tareas de recuerdo dependientes del hipocampo. Más aún, la administración de cortisol a niveles similares a los del estrés en sujetos normales, deteriora de forma selectiva la memoria verbal declarativa, sin afectar a la no-verbal (procedimental). Hay datos concordantes en pacientes con depresión e hipercortisolemia, o en pacientes con enfermedad de Cushing (secreción excesiva de corticosteroides).
La magnitud del estrés (en duración o intensidad) se refleja en el grado de afectación de la memoria y el aprendizaje, como puede verse en la figura 1.
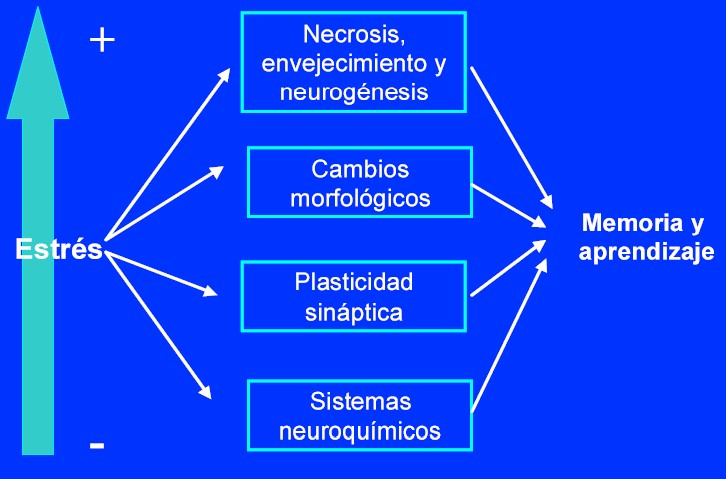
El estrés incontrolable facilita dos subtipos de condicionamiento de parpadeo2 en ratas. Resulta interesante observar que, a diferencia de este condicionamiento, en el condicionamiento al miedo en ratas (donde a su vez se diferenciarían dos subtipos, uno relacionado con el contexto que precisa del hipocampo, y otro independiente del contexto), el subtipo dependiente del hipocampo se afecta de manera selectiva en situaciones de estrés y con la administración de corticosterona. Así pues, parece que el estrés ejerce efectos complejos entre los distintos tipos de memoria del hipocampo. Puede ser relevante la generalización de que el estrés facilita el aprendizaje (relacionado con el miedo) dependiente del hipocampo (que en humanos denominaríamos memoria procedimental) y deteriora el procesamiento de la información espacial (el equivalente en humanos sería la memoria declarativa) que se adquiere del contexto en el que el condicionamiento aversivo se produjo.
Estrés y plasticidad en el hipocampo
En las tres últimas décadas, el modelo fisiológico primario de la memoria ha sido la potenciación a largo plazo (LTP) – un fortalecimiento sostenido de la eficacia sináptica que se produce tras una estimulación tetánica [consiste en un conjunto agrupado de estímulos (tren) que activan los axones aferentes brevemente a una frecuencia alta. En los experimentos de LTP, ésta se consigue habitualmente con un tren de 1 segundo de duración a una frecuencia de 100 Hz, es decir, 100 estímulos en un segundo] breve de las fibras aferentes excitatorias. Si esta hipótesis de que los cambios en la eficacia sináptica subyacen al almacenaje de la información (denominada postulado de Hebb) es válida, entonces el hallazgo de que el estrés deteriora la memoria dependiente del hipocampo, predeciría que el estrés interferiría asimismo con la inducción de la LTP hipocámpica (o procesos relacionados), lo cual ha sido extensamente comprobado en estudios in vivo e in vitro.
En el equivalente experimental de la indefensión aprendida (donde las ratas no pueden realizar ninguna respuesta adaptativa para escapar del estímulo aversivo), se produce un deterioro de la memoria y el aprendizaje. No obstante, si las ratas tienen algún control sobre la terminación del estímulo aversivo, muestran una LTP hipocámpica más robusta que en los controles, indicando que es ese aspecto psicológico incontrolable de la descarga lo que ejerce el efecto inhibidor primario de la LTP en el hipocampo.
Existe una forma de potenciación de sinapsis, denominada PBP que sería una forma de LTP con un umbral más bajo, y que tiene una mayor sensibilidad a la modulación por el envejecimiento, las drogas, las hormonas y la neuromodulación y, a su vez, al impacto sobre el procesamiento en el hipocampo de las variables comportamentales relevantes.
Además del deterioro de PBP y LTP, existe otro efecto producido por el estrés y la corticosterona en las sinapsis del hipocampo, una depresión homosináptica de larga duración en el área CA1 (LTD). Así pues, el efecto del estrés parecería afectar el rango fisiológico de la plasticidad sináptica de manera que se favorecería el desarrollo de la depresión sobre la potenciación. Estos efectos no son permanentes, sino que la modulación del funcionamiento del hipocampo por el estrés implicaría un proceso dinámico continuo bajo la influencia permanente de la percepción que se tiene del entorno.
Otros efectos del estrés en el hipocampo
Además de afectar a la memoria y a la plasticidad sináptica, el estrés y la corticosterona pueden alterar la morfología de las dendritas e inhibir la neurogénesis en el cerebro adulto, lo que potencialmente puede tener un impacto en la memoria.
-
reduzcan la transmisión de aminoácidos excitatorios (aspartato y glutamato) (p.eje. fenitoína)
-
reduzcan los niveles extracelulares de serotonina (5-HT) (p.eje. tianeptino)
-
reduzcan la excitabilidad general a través de una estimulación GABAérgica (p.ej. benzodiazepinas)
Estrés y morfología dendrítica
McEwen y cols han estudiado extensamente el tema, mostrando que el estrés o la administración de corticosterona sostenida (p.eje. 3 semanas) puede producir atrofia de las espinas dendríticas en diversas áreas del hipocampo (CA3, pero también CA1 y giro dentado). Dicha atrofia puede bloquearse con drogas que:
lo que habla de los múltiples neurotransmisores y sistemas hormonales implicados en la inducción de atrofia en las dendritas del hipocampo.
La participación de los receptores del NMDA (N-metil-D-Aspartato) es crucial y bien conocida en la plasticidad sináptica y la memoria. No obstante, la activación del receptor NMDA también parece ser un elemento central en la manifestación de los efectos adversos a corto y medio plazo en la morfología del hipocampo. Por ejemplo, en la muerte celular producida por la corticosterona se producen elevados niveles de Ca2+ que pueden ser reducidos con antagonistas del receptor NMDA. Resulta irónico que las células piramidales del hipocampo eliminen receptores NMDA para almacenar la información, aunque el mismo mecanismo, estimulado de forma crónica, puede abocarles a su eliminación.
Estrés y neurogénesis
La producción de células granulosas en el giro dentado del adulto está documentado en un amplio número de especies incluyendo la humana. Su significación funcional aún no se comprende del todo, pero parece relacionada con las demandas del aprendizaje y la memoria. Así por ejemplo, en situaciones de aprendizaje dependiente del hipocampo, su número total y la longevidad de las de nueva formación aumenta.
La producción de células granulosas se inhibe por el estrés, agudo o crónico, o con la administración experimental de corticosterona. Asimismo, la retirada de los corticosteroides endógenos produce la estimulación de la producción de nuevas células granulosas en ratas jóvenes y adultas [Hay que señalar que la verificación de la neurogénesis adulta está en cuestión por el artefacto que introducen las manipulaciones experimentales].
El mecanismo subyacente es desconocido, si bien dado que las células granulosas carecen de receptores para glucocorticoides (ni MRs ni GRs, ver más abajo), la influencia debe ser indirecta, quizás por medio de un incremento de la transmisión glutamatérgica. Al igual que el estrés afecta el aprendizaje dependiente del hipocampo y la potenciación de sinapsis a largo plazo (LTP), por medio de la liberación de glutamato y la activación del receptor NMDA, idéntico mecanismo inhibe la proliferación de células granulosas. Una vez más, el receptor NMDA sirve como un sitio de acción común para los efectos constructivos (aprendizaje y memoria) y destructivos (inhibición de la proliferación celular) de los glucocorticoides en el hipocampo.
Mediadores neuroquímicos de los efectos del estrés
El estrés desencadena un conjunto de respuestas neuroquímicas (glucocorticoides, catecolaminas, opioides...) del eje HHA y del sistema nervioso autónomo. Existen abundantes evidencias de que uno de los neuromoduladores principales de la potenciación de sinapsis a largo plazo (LTP) y la memoria es la corticosterona. Sus efectos en la plasticidad sináptica del hipocampo son complejos, pues tanto los niveles bajos (como en la adrenalectomía) como los elevados (en el estrés o en la administración exógena) se asocian con deterioro de la potenciación de sinapsis a largo plazo (LTP), mientras que el nivel máximo de potenciación sináptica ocurre con niveles intermedios de corticosterona. Esta respuesta bifásica se explica por la existencia de dos tipos de receptores, los mineralcorticoideos (MRs), que se activan a niveles bajos o intermedios de corticosterona y que aumentan la magnitud de la potenciación de sinapsis a largo plazo (LTP), y los glucocorticoideos (GRs) de baja afinidad, que se activan con niveles altos de corticosterona y que atenúan la potenciación y estimulan la depresión sináptica de larga duración (LTD). Así en condiciones normales, niveles bajos o intermedios de corticosterona activarían los MRs, potenciando la plasticidad sináptica en el hipocampo, mientras que en situaciones de estrés, los altos niveles de corticosterona resultarían en una inhibición de la plasticidad sináptica. La mayor importancia de los GRs en la mediación de los efectos adversos del estrés, se manifiesta en que los agonistas GR y los antagonistas MR pueden deteriorar la memoria, y que una mutación en el receptor GR, bloquea los efectos de la administración exógena de corticosterona en la memoria dependiente del hipocampo.
Sin embargo, el incremento de corticosterona per se no es suficiente para que se altere la potenciación de sinapsis a largo plazo (LTP) hipocámpica. Por un lado, la potenciación de sinapsis a largo plazo (LTP) puede inhibirse en situaciones de estrés en ratas adrenelectomizadas (sin corticosterona por tanto). Por otro, hay descritas 3 situaciones donde el aumento de los niveles de corticosterona en plasma no produjo cambios en la potenciación de sinapsis a largo plazo (LTP) hipocámpica: si existe una lesión en la amígdala; la administración exogena de corticosterona en situaciones no estresantes; y en ratas macho a las que se les daba acceso a hembras sexualmente receptivas. Así pues, en ausencia de estrés o sin una amígdala intacta, la elevación de la corticosterona no es suficiente para producir déficits en el procesamiento hipocámpico.
Otros neuromoduladores parecen tener un papel relevante en la mediación de los efectos del estrés en la plasticidad sináptica, como se deriva de los siguientes datos:
-
Péptidos opioides: la naltrexona (un antagonista opioide) bloquea los efectos del estrés en el aprendizaje y la potenciación de sinapsis a largo plazo (LTP).
-
Sistema serotonérgico: El estrés eleva la 5-HT en el hipocampo, y la administración exógena de 5-HT puede inhibir la potenciación de sinapsis a largo plazo (LTP) en el área CA1 in vivo e in vitro.
-
Receptor NMDA: los antagonistas NMDA bloquean los efectos del estrés en el aprendizaje y la potenciación de sinapsis a largo plazo (LTP), así como el deterioro mediado por los glucocorticoides de la potenciación de sinapsis a largo plazo (LTP).
Amígdala, estrés y funcionamiento del hipocampo
La amígdala desempeña un papel primordial para el aprendizaje emocional y para la manifestación de los efectos relacionados con el estrés tanto a nivel conductual como en el funcionamiento hipocámpico. La lesión o la supresión farmacológica de la amígdala previene los erosión gástrica, la analgesia, o la conducta ansiosa inducidos por el estrés, bloquea el efecto modulador de drogas sobre la memoria dependiente del hipocampo, y deteriora la potenciación de sinapsis a largo plazo (LTP) in vivo.
Los receptores NMDA de la amígdala parecen implicados en la regulación de la potenciación de sinapsis a largo plazo (LTP) hipocámpica. Así la infusión de antagonistas NMDA en el núcleo basolateral de la amígdala, deteriora la potenciación sináptica en el giro dentado (hipocampo). En cambio, la estimulación a alta frecuencia de ese mismo núcleo produce una potenciación de las sinapsis en el giro dentado. Más específicamente, la estimulación de la amígdala tiene un efecto bifásico dependiente del tiempo (inmediatamente excitatorio y uno inhibitorio más duradero) sobre la potenciación de sinapsis a largo plazo (LTP) del hipocampo.
Es interesante reseñar que las lesiones en la amígdala también previenen el incremento en la eliminación de catecolaminas inducido por el estrés, que se postula como mecanismo subyacente al deterioro de la memoria de trabajo en la corteza prefrontal.
Anatómicamente, la amígdala está conectada tanto directamente como indirectamente (a través de la corteza entorrinal) con diversas regiones del hipocampo.
La literatura disponible apoya de manera creciente que para la expresión completa de los efectos del estrés en el hipocampo se precisa de la coactivación de la amígdala y el hipocampo, junto con la acción directa de los neuromoduladores (corticosterona, 5-HT, opioides...) sobre el hipocampo.
Un modelo integrado
Los autores enfatizan que los efectos locales de los neuromoduladores actúan en conjunción con las influencias procedentes de la amígdala, para alterar la memoria dependiente del hipocampo y la plasticidad sináptica. Proponen un modelo que describe un subconjunto de interacciones neuroendocrinas, que son condiciones previas necesarias para los efectos del estrés en el funcionamiento del hipocampo. Este modelo parte de 5 premisas:
-
Las hormonas del estrés pueden afectar directamente al hipocampo.
-
Las hormonas del estrés pueden influir a la amígdala directa o indirectamente.
-
Las proyecciones amigdalares al hipocampo pueden modificar el funcionamiento del hipocampo.
-
El hipocampo (área CA1) puede funcionar normalmente sin la amígdala en situaciones no estresantes.
-
Para que se produzcan alteraciones en la plasticidad sináptica del hipocampo, se requiere del efecto conjunto de las proyecciones amigdalares y de las hormonas del estrés.
Las figuras 2 y 3 ejemplifican el modelo planteado por los autores. En condiciones normales (figura 2), frente a un estímulo emocional, y a través de las conexiones neurales de las vías perceptivas implicadas (vías neurales en la figura, señaladas mediante flechas de puntos), se activa un doble circuito: neural [conexiones sinápticas: en general de carácter ascendente en cuanto a la complejidad de la percepción (de subcortical a cortical, de áreas corticales sensoriales a áreas asociativas...) y bastante bien conocido para las distintas modalidades perceptivas] y endocrino, a través de la activación del eje Hipotálamo - Hipofiso - Adrenal (HHA), que implica al hipotálamo, la hipófisis y las glándulas suprarrenales [conexión a través de hormonas que viajan por el torrente circulatorio] (en la figura representado por flechas de línea continua)
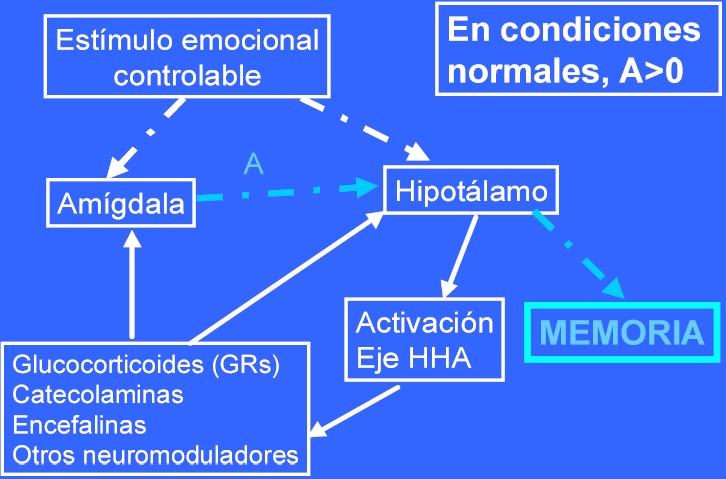
Si la amígdala está lesionada o se bloquea su activación (A=0) [inhibiendo o bloqueando los neuromoduladores intermediarios (corticoides, opioides, GABA, NMDA...)]
- Deterioro memoria/aprendizaje dependientes del hipocampo
Si se incrementa la actividad de la misma (A>>0) [idénticos neuromoduladores, pero mecanismo inverso]
- Potenciación de la memoria.
En situaciones de estrés (figura 3), la situación es inversa. La activación de la amígdala y su efecto modulador sobre el hipocampo (A>0) es determinante (unido al efecto de los neuromoduladores directamente sobre el hipocampo) para el deterioro de la memoria por el estrés. En aquellas situaciones (lesión de la amígdala) o cuando los neuromoduladores inhiben o bloquean la activación de la amígdala (A=0) –lo que en condiciones normales perjudicaría el aprendizaje/memoria dependientes del hipocampo- no se produce un deterioro de la memoria independientemente de la magnitud del estrés.
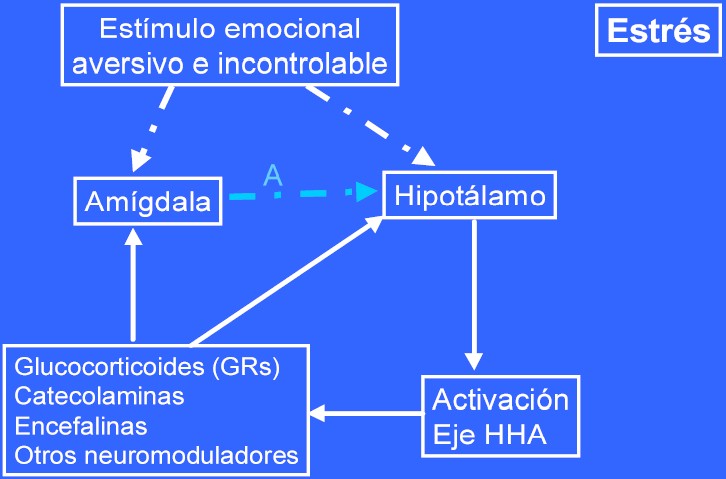
Es interesante comprobar como los neuromoduladores y receptores implicados son los mismos en ambas situaciones: potenciación de la memoria vs deterioro en la memoria secundario a estrés.
Notas
1.En realidad, la definición de estrés resulta una descripción nebulosa de diversas respuestas fisiológicas y conductuales. Una definición del fenómeno debería tener en cuenta los siguientes aspectos:
-
El que el estrés ocurra no depende de los parámetros físicos de la estimulación medioambiental, sino del modo en que el organismo percibe y reacciona a dicho estímulo.
-
No existe ningún estado fisiológico que sea específico del estrés. Por ejemplo, se acepta que una elevación del nivel de glucocorticoides es un indicador de un estado de estrés, pero estos también pueden elevarse en actividades placenteras, como la alimentación, el ejercicio o el sexo.
-
La controlabilidad del factor estresante tiene una influencia profunda en el impacto de una experiencia aversiva en la fisiología y la conducta. Así, numerosos estudios muestran que tener control sobre una experiencia aversiva mitiga profundamente la percepción de cuán estresante fue (como dato sobre el punto anterior, los niveles de glucocorticoides son casi insensibles a la controlabilidad).
Los autores proponen una definición con estos 3 componentes.
Primero, el estrés requiere una intensificación de la excitabilidad o el arousal, que puede ser medida operativamente empleando la electroencefalografía, la actividad conductual (motora) o mediante análisis neuroquímicos (adrenalina, glucocorticoides). Segundo, la experiencia debe ser percibida como aversiva, definiendo este hecho como una indicación de que el sujeto evitaría o atenuaría la intensidad del agente estresante si pudiera. Tercero, grado de controlabilidad de la experiencia, variable que determina en último término la magnitud de la experiencia estresante y de la susceptibilidad del individuo para desarrollar secuelas conductuales o fisiológicas relacionadas con el estrés.
2.El condicionamiento de parpadeo es del tipo pavloviano, en el que el sujeto aprende a responder con un parpadeo a un estímulo condicionado (EC; sonido o luz) que ha sido apareado de manera contingente con un estímulo no-condicionado (ENC; un chorro de aire o una descarga eléctrica al ojo). Se distinguen dos subtipos: el de retraso, donde el EC precede y se superpone al ENC; y el de trazo, donde existe un intervalo "vacío" entre el EC y el ENC. Ambos requieren la integridad del cerebelo pero, a diferencia del subtipo de retraso, en el de trazo se requiere además la integridad del hipocampo [sería un ejemplo de aprendizaje/memoria dependiente del hipocampo].
Bibliografía
Hebb, D.O. The organization of behaviour: a neuropsychological theory. Wiley, New York. 1949.
Kim, J.J. & Yoon, K.S. (1998) Stress: metaplastic effects in the hippocampus. Trends Neurosci.21, 505-9.
McEwen, B.S. (2000) Effects of adverse experiences for brain estructure and function. Biol Psychiatry 48, 721-31.
Newcomer, J.W., et al. (1994) Glucocorticoid-induced impairment in declarative memory performance in adult human. J. Neurosci. 14, 2047-53.
Sponsored Links : No Frills Flyer, Food Basics Flyer, Valu-Mart Flyer, Rexall Flyer, IGA Flyer