Número 018 2004 Revista Internacional de Psicoanálisis Aperturas
Resolviendo la dualidad crianza-ambiente en los orígenes de la ansiedad: princio de co-acción
Autor: de Iceta Ibáñez de Gauna, MarianoPalabras clave
Afectuosidad y ansiedad, Carga genetica de la ansiedad, Comorbilidad ansiedad y depresion, Crianza-ambiente en la ansiedad, Interpretacion de la experiencia, Modulacion del transporte de serotonina, Tamaño del hipocampo..
La ansiedad es un estado mental que se desencadena como anticipación de una amenaza real o potencial. Forma parte de las respuestas fisiológicas en humanos y encontramos respuestas equivalentes en otras especies.
No obstante, como cualquier terapeuta conoce, cuando se vuelve excesiva o inapropiada puede convertirse en una enfermedad. Dentro de los diagnósticos categoriales, como por ejemplo el DSM-IV, se subclasifica en seis trastornos según el grupo de síntomas que aparezcan asociados (Ansiedad Generalizada, Trastorno de pánico, Trastorno por Estrés Postraumático, Fobia social, Fobia específica, y Trastorno Obsesivo-compulsivo). Igualmente, aparece como un síntoma en muchas otras entidades nosológicas, desde los trastornos del estado del ánimo, a la esquizofrenia.
Desde que existe una teoría de la enfermedad, viene transcurriendo el debate entre los defensores del origen biológico/genético (1) , y los que ubican el origen de la patología en la crianza/ambiente, posición esta última en la que tradicionalmente se ha alineado el psicoanálisis.
Este debate ha sido general en torno a todos los trastornos mentales, pero en algunos casos, como la esquizofrenia, la mayor evidencia de factores genéticos o biológicos ha suavizado el debate llevándolo a modelos donde la genética/biología aportaría una cierta vulnerabilidad y el ambiente el elemento desencadenante para la aparición última del trastorno; en el caso de los trastornos de ansiedad, por la menor contribución de la genética, las posiciones han permanecido firmes.
Es necesario precisar que no obstante, en algunos trastornos, unas posiciones se mantenían más fuertes que en otros. Así, por ejemplo, numerosos estudios relacionan determinadas características físicas como por ejemplo la laxitud articular (hiperflexibilidad), o la respuesta a la inhalación de CO2 con el trastorno de pánico, mientras que en el caso de la ansiedad generalizada o la fobia, los orígenes ambientales o en la crianza pueden encontrar más defensores. Planteando implícitamente subclasificaciones como trastornos de ansiedad más “biológicos” y otros más “ambientales”.
Vamos a repasar algunos aportes en los últimos meses en este sentido, con datos suficientes como para resolver la dicotomía del debate:
Una reciente revisión de Gross y Hen (Gross & Hen, 2004) contribuye a deshacer este debate dicotómico planteando la ansiedad como el resultado de experiencias ambientales precoces, cuyo impacto está modulado por factores de susceptibilidad genéticos.
En su trabajo abordan cuatro apartados: la fisiología de la ansiedad, los mecanismos moleculares implicados, las interacciones genética-ambiente, y la influencia de eventos en el desarrollo en la ansiedad adulta. Por su interés para nuestro trabajo como terapeutas, me centraré en estos dos últimos aspectos.
Interacciones genética-ambiente y ansiedad
Las personas poseemos un rasgo bastante estable a lo largo de la vida, un cierto nivel de ansiedad. Esta diferencia en el grado de conductas ansiosas persistente en el tiempo que presentamos, es un reflejo de diferencias estructurales o de función en los cerebros de los individuos muy ansiosos frente a los poco ansiosos. Tales diferencias se desarrollan como resultado de las diferencias en cuanto a su bagaje genético y las experiencias a lo largo de la vida.
Los estudios de gemelos confirman dicha hipótesis. Cuando se estudia la aparición de trastornos de ansiedad en gemelos monocigóticos (comparten exactamente la carga genética) con gemelos dicigóticos, se demuestra que sólo se puede atribuir a la genética un 30-40% de la responsabilidad en el origen de dichos trastornos. Este porcentaje es relativamente bajo cuando se compara con el de otras enfermedades psiquiátricas como la esquizofrenia (60-70%) o neurológicas como la enfermedad de Huntington (100%).
Estimar las diferencias ambientales resulta mucho más difícil. Uno de los elementos son los estudios de adopción. En ellos se asume que los gemelos criados juntos tienen el mismo ambiente y se compara con los casos donde fueron adoptados por familias diferentes. Inesperadamente, el resultado sólo atribuye al ambiente un 5% de la variación en la aparición de un trastorno. Este bajo resultado puede deberse a que los gemelos experimenten el ambiente compartido de forma diferente. A su vez las experiencias individuales o compartidas son probablemente modificadas por, o dependientes de, factores genéticos (interacciones genética-ambiente) o el producto de los mismos (correlaciones genética-ambiente). En los trastornos como la ansiedad, donde la contribución genética es modesta, estas interacciones y correlaciones son seguramente de mayor importancia.
En humanos sólo se han encontrado unas pocas variaciones genéticas ligadas a un aumento de la ansiedad. Diversos estudios han mostrado un pequeño pero significativo incremento de la ansiedad en niños y adultos que poseen una variedad en el gen del transportador de la serotonina. La distribución de esta variación genética en la población es: 32% tienen los dos alelos, homocigotos (baja actividad del transportador de serotonina, alto nivel de neuroticismo); 49% sólo tienen uno heterocigotos; 19% no la tienen en ninguno de sus dos alelos. En estudios recientes con resonancia magnética funcional han mostrado que la presencia de este rasgo se asocia con un incremento de la actividad de la amígdala ante la presentación de caras asustadas. Esto es, el transportador de serotonina influye la conducta relacionada con la ansiedad modulando la excitabilidad de circuitos del miedo específicos en el cerebro. Dichos resultados parecen discordantes con la eficacia terapéutica de los ISRS, que bloquean la acción del transportador de serotonina. No obstante, encuentran apoyo en estudios con ratones donde se desactiva este gen produciéndose un aumento de la ansiedad.
Resulta intrigante el hecho de que el bloqueo farmacológico del trasportador de serotonina (análogo a los ISRS) en las dos primeras semanas de vida produce un fenotipo (la expresión final de una determinada carga genética o genotipo) con incremento de ansiedad (como los homocigotos). Así pues, parece que la modulación del transporte de la serotonina tiene efectos opuestos en la regulación de las conductas relacionadas con la ansiedad en el desarrollo y en la edad adulta.
El trastorno por estrés postraumático (TEP, de desgraciada actualidad en Madrid) es un ejemplo donde los factores de riesgo ambientales parecen ser regulados por factores genéticos. Aproximadamente, el 15% de las personas que experimentan o son testigos de un trauma severo desarrollarán un TEP. Uno de los hallazgos más consistentes es la reducción del tamaño del hipocampo (2) en los pacientes con TEP. Existen datos acerca de la facilidad con que las hormonas del estrés (Ver Neurobiología de las interacciones estrés memoria, en el No. 11 de Aperturas, para mayor detalle) lesionan el hipocampo, y diversos autores han propuesto que la disminución de tamaño de esta región cerebral en los pacientes con TEP es consecuencia directa del estado de estrés crónico que induce el trauma.
Sin embargo, estudios de neuroimagen recientes de gemelos discordantes para el TEP (3) indican que esta hipótesis es incorrecta. Así, compararon los cerebros de 40 parejas de gemelos monocigóticos (idénticos); en todos los casos, uno de los gemelos fue a la guerra de Vietnam y entró en combate, mientras que el otro se quedó en casa. De los que combatieron, un 42% desarrollaron TEP. Sin embargo, al comparar sus cerebros con los de sus gemelos que no sufrieron el trauma, no encontraron diferencias significativas (en ambos casos tenían hipocampos de volumen reducido), luego la reducción de volumen no era consecuencia del trauma sino una condición pre-existente. Más interesante aún es que encontraron una correlación inversa entre el tamaño del hipocampo y el riesgo de padecer TEP, lo que podría explicar el motivo por el que sólo algunos de los que experimentan el trauma desarrollan TEP, indicando que el tener un hipocampo pequeño aumenta la susceptibilidad individual al estrés ambiental.
Queda por dilucidar la contribución relativa al tamaño del hipocampo, ¿es el ambiente o la genética lo que determina este rasgo que condiciona una mayor susceptibilidad al estrés? Una vez más, la respuesta parece ser ambos factores son determinantes. Así, los datos actuales procedentes de estudios en roedores y en humanos, apoyan un origen mixto, donde los factores genéticos son importantes, y la influencia del ambiente sería mayor en el desarrollo (desde mitad de la gestación a la adolescencia) que en etapas posteriores.
Acontecimientos durante el desarrollo y Ansiedad en adultos
Existe abundante información que apoya el hecho de que la susceptibilidad a la psicopatología puede determinarse tempranamente en la vida. No obstante, así como el padecer un trauma incrementa el riesgo de patología en la edad adulta, no existe especificidad. Esto es, ningún trauma concreto determina una patología concreta en la edad adulta, lo que indica que otros factores (posiblemente genéticos) determinan la patología precisa que es desencadenada por el trauma infantil.
Dos estudios longitudinales recientes sobre niños expuestos a un entorno familiar violento constituyen un ejemplo llamativo de esta interacción.
En el primero, el maltrato severo a edad temprana se asociaba con un incremento significativo del riesgo de conductas antisociales en la adolescencia y la edad adulta, incluyendo trastornos de conducta, tendencia a ser violento y el haber sido declarado culpable de una agresión. Hasta aquí, algo conocido. Lo interesante es que el grupo de Caspi encontró que el impacto del maltrato se modificaba de manera muy importante en función de un determinado rasgo genético (el promotor del gen que codifica la actividad MAO, una enzima que se encarga de metabolizar la serotonina, la dopamina y la noradrenalina). En aquellos niños con un gen de baja actividad, el maltrato constituía un factor de riesgo, mientras que cuando poseían el gen de alta actividad el maltrato no suponía diferencia en términos de conducta antisocial en la edad adulta. Esto implica que las consecuencias bioquímicas de una actividad MAO alta son suficientes para proteger al cerebro de las consecuencias a largo plazo (en cuanto a conducta antisocial) del abuso en la infancia.
En un segundo estudio del mismo grupo (Caspi et al, 2003) en la misma muestra, encontraron una relación entre la depresión en la edad adulta y el haber padecido abusos en la infancia o el número de eventos estresantes durante la misma. Lo interesante, al igual que en el otro estudio, es que esta asociación se daba sólo en aquellos niños con una determinada configuración genética, mientras que no tenía ese efecto en los que no la tenían (en este caso, los que poseían genes que codifican una alta actividad del transportador de la serotonina [5-HTT] parecen estar protegidos frente al efecto predisponente a la depresión de los sucesos estresantes). Cabe destacar asimismo que la predisposición a la depresión no se modificaba por las diferentes configuraciones genéticas (polimorfismo) de la actividad MAO, lo que indica que ambas susceptibilidades tienen diferentes mecanismos moleculares mediadores. Dada la alta comorbilidad (4) de la ansiedad y la depresión, y las evidencias de su modulación por factores genéticos comunes (revisión de Kendler, 1996), es probable que la predisposición a los trastornos de ansiedad esté igualmente determinada por influencias durante el desarrollo cuyo impacto en el cerebro se encuentre regulado genéticamente.
La observación de que los individuos son particularmente susceptibles a las influencias ambientales adversas fue confirmada por estudios en animales que mostraron los efectos poderosos que la calidad del cuidado materno tenía en la conducta emocional y el funcionamiento cerebral a lo largo de la vida.
- En la cría de mono rhesus, la sustitución durante los primeros meses de vida de la madre por un objeto inanimado induce deficiencias a largo plazo en la interacción con iguales y la adaptación social. También se asocia con un incremento del riesgo de desarrollar conductas relacionadas con la ansiedad, como el balanceo y el acicalamiento.
- En una raza de macacos, el aumento de la impredecibilidad en la búsqueda de comida para las madres hace que las crías tengan alteradas la hormona del estrés y la respuesta al miedo cuando sean adultos.
- En los monos rhesus, como en humanos, existe un polimorfismo del transportador de la serotonina (5-HTT). Al igual que en humanos, una de las configuraciones (el alelo corto) se asocia con un aumento de las conductas relacionadas con la ansiedad y del metabolito de la serotonina (5-hidroxi-indol-acético, 5-HIAA). Resulta intrigante que en los monos el efecto del polimorfismo 5-HTT está regulado de manera poderosa por el ambiente de la crianza temprana. Así, los monos criados por sus madres tienen niveles normales de 5-HIAA independientemente de su configuración genética. En cambio, los criados en grupos desde los 30 días a los 7 meses presentan niveles altos (alelo corto) o bajos (alelos largos) de 5-HIAA en la edad adulta. Estos datos indican que el impacto fisiológico del polimorfismo 5-HTT depende de las interacciones maternas y sociales precoces.
- En roedores se han realizado numerosos estudios con resultados similares:
- Las ratas que fueron separadas de sus madres en el período postnatal precoz durante varias horas al día muestran conductas relacionadas con la ansiedad y un aumento de la reactividad hormonal al estrés.
- Las ratitas criadas por madres con déficits en las conductas de lameteo y acicalamiento (“poco mimosas”) tienen mayores niveles de comportamientos relacionados con la ansiedad que las que tuvieron madres con altos niveles de las mismas conductas (“muy mimosas”). Los estudios de crianza cruzada mostraron que dichas influencias eran primariamente ambientales (Ver Figura 1).
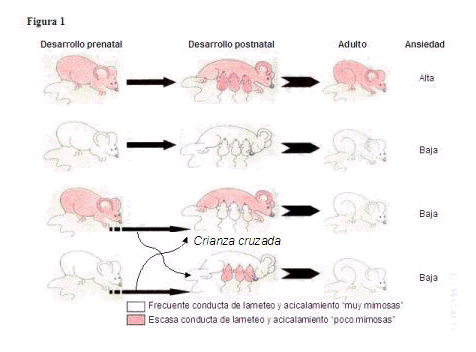
Cuando se intercambiaban las crías de las ratas “poco mimosas” a una “muy mimosa” disminuía el riesgo de desarrollar comportamientos relacionados con la ansiedad en la edad adulta. Lo inverso, en cambio, no se daba. Las crías de madres “muy mimosas” criadas por una madre “poco mimosa” no tenían un mayor riesgo de tener ansiedad al llegar a ser adultos. Esto quiere decir que existen factores genéticos o del ambiente intrauterino de las madres “muy mimosas” que confieren protección contra los efectos adversos de la crianza posterior. El trasplante de embriones, casi inmediatamente tras la concepción, de ratas “muy mimosas” al útero de ratas “poco mimosas”, unida a la crianza sí era suficiente para producir una camada “poco mimosa”. Así pues, hay señales pre- y post-natales de las madres que actuando sinérgicamente pueden inducir cambios a largo plazo funcionales y estructurales en los circuitos de la ansiedad.
- Por último, se ha mostrado que la conducta “muy mimosa” adquirida experimentalmente, puede transmitirse de una generación a otra. Las ratas hembra criadas por madres “muy mimosas” lo serán con sus crías, y tendrán camadas con poca ansiedad, independientemente de que su madre biológica fuera de las “poco” o “muy mimosas”. Esta herencia epigenética de las conductas relacionadas con la ansiedad subraya la influencia que los factores ambientales pueden ejercer en circuitos cerebrales remodelados de forma persistente durante el período temprano del desarrollo.
En la línea de lo planteado por los autores de esta revisión, el grupo de Michael Meaney ha publicado en Nature Neuroscience el posible mecanismo a nivel molecular para explicar cómo la conducta materna puede producir cambios a largo plazo en la prole. Nuevamente se evidencia el pricipio de co-acción para este efecto, y la existencia de un período ventana precoz en el desarrollo para producir estos cambios moleculares con repercusión en la regulación del estrés a largo plazo (Weaver et al, 2004).
Por último, quisiera recuperar un reciente trabajo publicado en Aperturas No. 17 por Emilce Dio Bleichmar (Modelos interactivos entre la genética de la conducta y la parentalización), ya que aborda el tema que nos ocupa en cuanto al debate sobre la influencia de la genética vs la parentalización (5) en el desarrollo.
El principal concepto que Dio Bleichmar destaca es el de la co-acción entre genética y ambiente:
“El concepto llave es entender que lo que hace que el desarrollo suceda es la relación entre los dos componentes, y no los componentes en sí (persona-persona, organismo-organismo, organismo-ambiente, célula-célula, gen-gen. actividad-conducta motora). Cuando se habla de co-acción como el corazón de la causalidad evolutiva lo que se quiere enfatizar es que necesitamos especificar alguna relación entre al menos dos componentes del sistema del desarrollo. El concepto usado más frecuentemente para designar la co-acción es el relativo a la experiencia”.
Repasa a continuación distintos estudios mostrando la influencia del medio externo:
“Es decir, si la corteza cerebral de una determinada zona recibe estímulos de un tipo dado, son estos estímulos los que ocasionan la organización y la funcionalidad que tendrá y no la predisposición genética. En otras palabras, es la naturaleza del estímulo externo el factor decisivo de cómo se terminará organizando una cierta zona cortical” [negrita en el original]
Destaca el énfasis en la interacción de ambos factores, lo genético y lo ambiental, así por ejemplo en relación con el desarrollo de un fenotipo agresivo en ratones
“ni un genotipo particular ni un factor traumático externo están inevitablemente vinculados a un resultado patológico. Es la particular combinación de factores genéticos y ambientales y probablemente el factor temporal de la interacción que provoca resultados patológicos” [negrita en el original].
Uniendo lo anterior al concepto de plasticidad cerebral concluye:
“la idea cada vez más fundamentada que en humanos hay un período en que el cerebro es plástico, durante el cual se organiza la funcionalidad y que, superada esa ‘ventana’ durante la cual el cerebro se puede modificar en una dirección determinada por el estímulo externo, después ya resulta mucho menos factible” [negrita en el original].
Más adelante, recoge las palabras de Kandel (6) quien introduce el papel de lo subjetivo en la vivencia de lo ambiental:
“El que un factor ambiental desencadene o no la expresión de un gen puede depender del modo en que el individuo interprete esa experiencia, determinada a su vez por significados conscientes o inconscientes atribuidos a la misma (Kandel, 1998). Así, los procesos representacionales intrapsíquicos no son sólo consecuencia de los efectos ambientales y genéticos, sino que es probable que sean moduladores vitales de dichos efectos”.
El mismo autor llegó a sugerir que los cambios más profundos producidos por la psicoterapia tendrían lugar a través de este mecanismo de modulación genética.
Quisiera incluir por último un esquema que la autora reproduce porque el mismo explica muy bien las influencias bidireccionales a múltiples niveles y en constante interacción en cuanto a la causalidad de los fenómenos del desarrollo y por ende (con el matiz de las ventanas de mayor plasticidad cerebral) del proceso de la psicoterapia en cuanto al cambio.
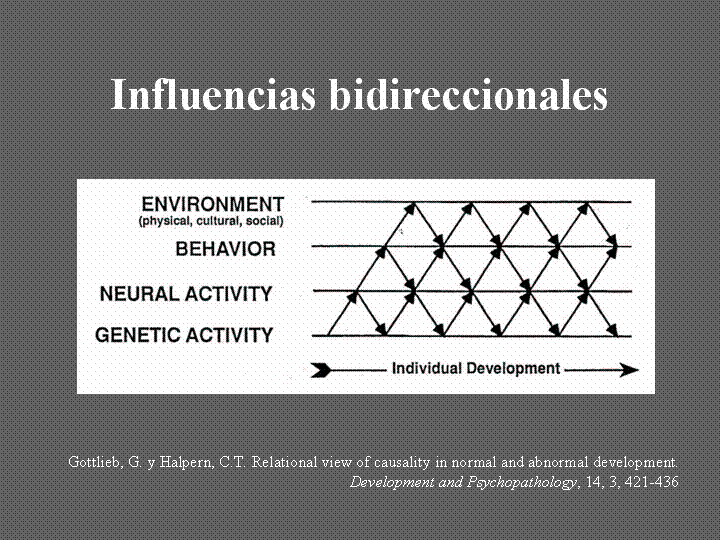
Todo lo anterior refuerza la necesidad de modelos complejos, como el enfoque modular-transformacional, capaces de abordar las complejidades de dichas interacciones tanto en el desarrollo de la teoría como de instrumentos técnicos precisos para producir el cambio terapéutico.
NOTAS
(1) Existen matices entre ambas posiciones, pero en cuanto oposición a la experiencia/ambiente como origen de la patología, las agrupamos conjuntamente.
(2) El hipocampo es una estructura ubicada en la cara medial del lóbulo temporal del cerebro que se requiere para la memoria asociativa.
(3) Uno de los gemelos lo padece y el otro no.
(4) Aparición conjunta de dos o más enfermedades, no implica relación causal entre las mismas.
(5) Lo cual en un sentido más amplio, puede referirse al propio proceso de la psicoterapia psicoanalítica, que en muchos casos constituye una segunda parentalización
(6) Premio Nobel 2000
Bibliografía
Caspi, A. et al. (2002) Role of the genotype in the cicle of violence in maltreated children. Science, 297: 851-854.
Caspi, A. et al. (2003) Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science, 301, 386-389.
Dio Bleichmar, E. (2004) Modelos interactivos entre la genética de la conducta y la parentalización. Revista internacional Aperturas Psicoanalíticas nº 17, http://www.aperturas.org.
Gross, C., Hen, R. (2004) The developmental origins of anxiety. Nature Reviews Neuroscience, 5: 7, 545-552.
Kandel, E.R. (1998) A new intellectual framework for psychiatry. Amer. J. Psychiat. 155: 457-69.
Kendler, K.S. (1996) Major depression and generalised anxiety disorder. Same genes, (partly) different enviroments ¾ revisited. Br. J. Psychiatry (Suppl.) 68-75.
Weaver, I.C.G. et al. Epigenetic programming by maternal behavior. Nature Neuroscience, 27 Jun 2004.